
 Аннотация. Настоящая работа рассматривает на примере биологических, социальных и технико-информационных систем популярное в современном научном сообществе понятие сетевые структуры (сети). Отмечается, что данное понятие применяется в двух различных значениях: 1) в широком смысле – как любая система из узлов (вершин), соединенных рёбрами (дугами); в таком значении понятие «сеть» синонимично понятию «граф»; 2) в более узком смысле – как децентрализованная кооперативная система, противопоставляемая системам с единым центром (лидером, пейсмейкером), т.е. иерархиям и в то же время системам, взаимоотношения между элементами которой носят в основном конкурентный, а не кооперативный характер – рынкам и их аналогам (квазирыночным структурам).
Аннотация. Настоящая работа рассматривает на примере биологических, социальных и технико-информационных систем популярное в современном научном сообществе понятие сетевые структуры (сети). Отмечается, что данное понятие применяется в двух различных значениях: 1) в широком смысле – как любая система из узлов (вершин), соединенных рёбрами (дугами); в таком значении понятие «сеть» синонимично понятию «граф»; 2) в более узком смысле – как децентрализованная кооперативная система, противопоставляемая системам с единым центром (лидером, пейсмейкером), т.е. иерархиям и в то же время системам, взаимоотношения между элементами которой носят в основном конкурентный, а не кооперативный характер – рынкам и их аналогам (квазирыночным структурам).
Сети в узком смысле в свою очередь могут быть подразделены на конкретные организационные варианты. Из числа этих вариантов в настоящей работе рассмотрены клеточные, модульные, эквипотенциальные, нейронные, эусоциальные, эгалитарные сети. В работе продемонстрировано, как количественные критерии, разработанные в рамках современной «network science», могут быть использованы для разграничения структур по линии сети-иерархии-(квази)рынки. В частности, иерархические и сетевые структуры (плоские и объёмные) могут быть отделены друг от друга по критериям совпадения-несовпадения разных критериев центральности их элементов (узлов). Сети и (квази)рынки могут быть разграничены с применением экологических плюс- и минус-критериев; дополнительное подспорье в этом плане даёт такая характеристика сетей как коэффициент кластеризации.
ВВЕДЕНИЕ
Сетевые структуры (сети) стали модным понятием в настоящее время. Сетевые структуры воплощены отнюдь не только в виртуальном мире Интернета и WorldWide Web, с которыми это понятие ассоциируется у многих читателей. В то же время сети – это не только приспособление для ловли рыбы, воспетое в сказках А. С. Пушкина. Сетевые структуры представляют собой междисциплинарное понятие, применимое к разнообразным системам — от звёздных скоплений и кристаллов до ансамблей элементарных частиц. Впрочем, в этой работе мы ограничиваемся лишь биосистемами, человеческим обществом и техническими устройствами (сравнивая эти три типа систем). Данная работа продолжает серию публикаций о сетевых структурах [13-19] и существенно дополняет предшествующие работы, где ставился вопрос о математических критериях классификации сетевых структур [20, 21].
Однако, как можно определить понятие «сетевая структура»? В литературе данное понятие применяется в по крайней мере двух различных значениях.
ШИРОКИЙ СМЫСЛ ПОНЯТИЯ «СЕТЕВАЯ СТРУКТУРА»
Сетевые структуры в более широком смысле могут быть определены как любые системы из элементов (вершин, или узлов) со связями между ними, которые обозначаются как ребра (дуги) [71]. Данное определение принимается многими учёными во всём мире.
Понятие «сеть» применялось в гуманитарных науках для описания отношений между индивидами (группами) в социуме примерно с начала XX века. Георг Зиммель, основатель «формальной школы» в социальных науках, использовал понятие «социальная сеть» в своих работах [83]. По словам Леопольда фон Визе, представителя этой школы, если бы мы могли остановить «постоянно струящийся поток межчеловеческих взаимодействий», то «мы бы тогда увидели кажущуюся непроницаемой сеть линий между людьми» [88]. Антрополог А. Р. Рэдклифф-Браун писал, что «прямое наблюдение показывает нам, что… человеческие индивиды соединены сложной сетью социальных отношений. Я использую термин „социальная структура“ для обозначения этой сети…» [78]. В своём президентском послании к английским антропологам в1940 г. Рэдклифф-Браун призывал к систематическому изучению сетей.
В 30-40-е годы прошлого века психиатр Джакоб Морено и психолог Хелен Дженнингс изображали в форме сетевой структуры взаимоотношения между индивидами в своих пионерских исследованиях неформального лидерства в американской тюрьме Синг-Синг и Гудзоновской школе для девушек [59, 69]. В1939 г. было проведено и в1941 г. опубликовано исследование (The “Southern Women Study”) над 18 женщинами южных штатов США, которые в целом посетили 14 социальных мероприятий. Авторы изобразили в форме сети диаграмму, в которой показано, какие из женщин участвовали в каких мероприятиях [48]. Тем не менее, в строго определённом значении термин «сеть» (network) стал использоваться в социологии лишь спустя полтора десятилетия, начиная с работ Дж. А. Барнеса [34].
Понятие «сетевые структуры» в такой интерпретации применялось к разнообразным типам систем [21, 30, 33, 53, 71-73, 80, 91]. Оно достаточно популярно в социологии, социальной психологии и ряде других научных областей, включая активно развивающуюся ныне «сетевую науку» (network science) [33, 71-73]. В человеческом социуме можно привести многочисленные примеры сетевых структур, в том числе: семья, где члены семьи служат точками (узлами), а соединяющими линиями (рёбрами) — их взаимоотношения; любая организация, рассмотренная с точки зрения взаимосвязей (рёбер сети) между ее работниками или их группами (узлами сети); несколько коммерческих предприятий, связанных между собой контрактными отношениями; социальное движение, объединённое некоторой идеей и многое другое. На макроуровне социума сетями являются разветвлённые транспортные комуникации, финансовые организации с множеством филиалов, переплетение городских систем жизнеобеспечения, а также, конечно, Интернет.
Подчеркнём, что сетевые структуры в широком смысле формируются также и объектами, не относящимися к человеческому социуму. Многие соответствующие примеры относятся к области биологии. Сетевой подход начали активно применять в последние годы (см.[57, 71, 91] ) к сообществам животных. Есть работы, где понятие сети в широком смысле прилагается, например, к исследованию стай рыб [44-46], групп летучих мышей [76], сурикатов [66] или шимпанзе [64]). Сетевой подход использовали при описании отношений доминирования-подчинения в централизованных сетях [31, 32, 49], которые мы обозначаем как иерархии в дальнейшем тексте этой статьи. Здесь необходимо отметить исследования нервной системы и особенно головного мозга, где широко применялось понятие «нервные, или нейронные, сети». Под нейронными сетями, начиная с пионерских работ У. С. Мак-Каллока и В. Питтса [67], понимали любые группы взаимодействующих между собой нервных клеток, рассмотренные вместе со связями между ними.
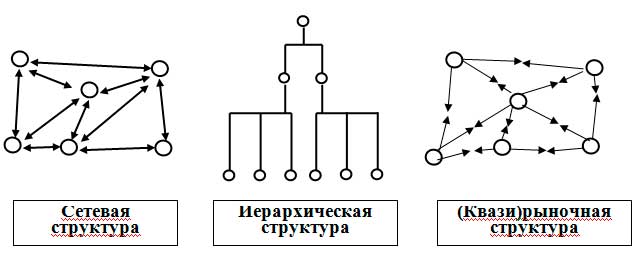
Рис. 1. Схематическое изображение сетевой, иерархической и (квази)рыночной структур; ← → кооперативные; → ← конкурентные взаимодействия элементов структур.
УЗКИЙ СМЫСЛ ПОНЯТИЯ «СЕТЕВАЯ СТРУКТУРА»
Начиная приблизительно c 1980-х годов, в литературе, в основном посвящённой наукам о человеке и обществе, была использована иная интерпретация термина «сетевые структуры» [35, 40, 42, 43, 60, 61, 68, 85]. В соответствие с ней, далеко не все системы из соединённых связями элементов следует именовать «сетевыми структурами».
В сети должен отсутствовать единый центр (лидер, доминант), и ее поведение является результатом кооперативных взаимодействий между элементами (узлами), среди которых могут быть несколько частичных лидеров с ограниченным воздействием на систему. Следует отметить, что всемирная паутина устроена во многом именно по данному организационному принципу. В настоящей работе термин «сетевая структура» будет употребляться в узком смысле, если не оговорено обратное.
Опираясь на целый ряд классических работ и публикаций последних лет [5, 6, 28, 35, 40, 42, 43, 60, 61, 68, 85], мы будем сопоставлять сетевые структуры с:
Сетевые структуры в человеческом социуме. В социуме сетевые структуры представляют собой «форму спонтанного порядка, которая возникает в результате действий децентрализованных агентов, а не создаётся какой-либо центральной властью» [27]. Предпосылками сетевых структур в социуме являются репутации их членов, их дружеские отношения, взаимозависимость и кажущийся непрагматичным альтруизм, который консолидирует сеть. Именно такие отношения обозначаются как социальный капитал» в социологии и теории менеджмента. Кооперация между членами сети стимулируется:
Сетевые структуры представляют собой «третью альтернативу вертикальному планированию», типичному для централизованной иерархии, а также «анархии рынка» [68]. Принципы сетевой организации могут быть реализованы коммерческими предприятиями, научно-исследовательскими лабораториями, социальными движениями и политическими организациями. Как всякая новаторская технология (будь то, скажем. ядерная энергетика или генетическая инженерия), сетевые структуры могут вызывать как положительные, так и отрицательные последствия. Угрозу современному миру представляют, например. «тёмные сети» наркодилеров, гангстеров или террористов. Хотя подобные структуры, включая Аль-Каеду, не имеют централизованной иерархической структуры, они включают влиятельных и потенциально устранимых частичных лидеров (то, что в «сетевой науке» обозначается как «хабы»).
Независимо от целей и конкретных норм поведения, сетевые децентрализованные кооперативные структуры в социуме не препятствуют конкуренции (соревнованию) между участниками в той мере, в какой конкуренция способствует креативности и целеустремлённости в решении стоящих перед всей сетью задач (проблем); однако, кооперация внутри сети преобладает над конкуренцией. Многие сетевые структуры следуют лозунгу «конкуренция ради успешной кооперации». Например, сетевая творческая лаборатория реализует научно-иследовательский или технологический проект, и модераторы (психологические лидеры) этой сетевой структуры направляют естественную конкурентную установку участников проекта в сторону ускорения работы по достижению общесетевой цели, скажем, разработки нового лекарственного препарата.
Социальные сети можно проиллюстрировать примером структуры под названием хирама (hirama). Это сокращение означает High-Intensity Research and Management Association ([14-16, 18-21, 74, 75]. Речь идёт о креативной команде (рис. 2), создаваемой для решения междисциплинарной задачи, например, Исследование локальных загрязнителей окружающей среды или Организованный терроризм и борьба с ним.
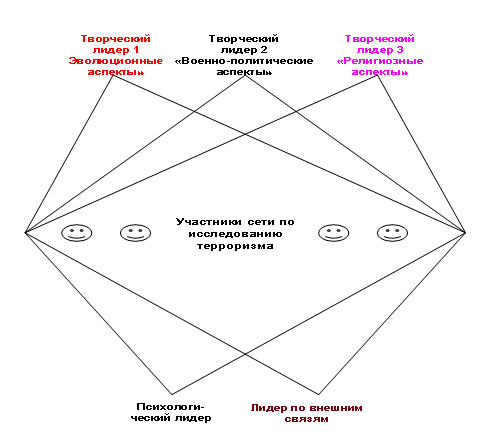
Рис. 2. Сетевая структура типа хирамы, нацеленная на различные аспекты исследования международного терроризма с целью борьбы с ним.
Задача дробится на несколько субпроблем, например, отмеченная задача борьбы с терроризмом может быть подразделена следующим образом:
Однако членение проблемы на субпроблемы не означает деление коллектива участников на части. Они параллельно работают по нескольким субпроблемам сразу — выступают не как узкие специалисты, а как генералисты (люди с гибкими функциями, переменной специализацией). За каждой из субпроблем закреплен только специализированный частичный творческий лидер, координирующий работу всех участников хирамы по соответствующей теме и протоколирующий их идеи. В помощь этому частичному лидеру могут быть приданы один или несколько экспертов, специалистов по профилю ведомой лидером субпроблемы. Специализированный частичный лидер и помощники-эксперты взаимодействуют с неспециализированными членами сетевой структуры хирамы, которые во многих хирамах численно преобладают.
В хираме имеется также психологический лидер, призванный налаживать отношения между индивидами и группами в коллективе и направлять их в конструктивное русло, смягчать конфликты, способствовать успешной работе по всем подпроблемам. Структура может включать также лидера по внешним связям (внешнего лидера), представляющего данную сетевую структуру в социуме, координирующего контакты с другими организациями и озвучивающего те или иные «наказы», петиции и др. документы, выработанные всем коллективом хирамы. Организационный лидер особенно важен на начальном этапе, когда сетевая структура организует свою деятельность и приобретает легальный статус (например, оформляется как организация по законам Российской Федерации). Возможны другие частичные лидеры, в зависимости от специализации данной хирамы.
Необходимо подчеркнуть, что по шаблону хирамы, с теми или иными вариациями, были построены междисциплинарные научные группы (например, биотехнологический центр DNAX в Калифорнии), коммерческие предприятия (компания Semco в Бразилии), а также разнообразные коммунарские и общинные структуры.
Сетевые структуры в биосистемах. Как отмечено в начале настоящей работы, термин «сетевые структуры» в широком смысле (любая система, состоящая из узлов и связей между ними), применялся к биологическим системам в целом ряде недавних публикаций. Возникает вопрос, приложим ли к биосистемам также и термин «сети» в узком смысле?
Представляется очевидным, что значительное число разнообразных биосистем имеет децентрализованный характер и в то же время характеризуется преобладанием кооперации элементов над конкуренцией между ними. В колонии кишечнополостных нет единого управляющего центра, и речь идёт о «нецентрализованной регуляции» их поведения [12]. Сетевые децентрализованные структуры весьма характерны и для мира микроорганизмов. Например, колонии микроорганизмов или их биоплёнки состоят из множества микробных клеток и устроены так, что отсутствие единого управляющего центра не препятствует эффективной координации социального поведения [17, 29, 35-37, 81, 82]. Клетки в составе многих тканей организма животных также организованы по принципам сетевой организации.
Можно привести немало примеров биосистем, для которых понятие «сетевые структуры» имеет не только чисто организационный, но и сугубо геометрический смысл. Хищные диктиобактерии образуют ловчие сети из множества бактериальных клеток; добыча застревает в ячейках этих сетей, несколько напоминая клиентов, «застревающих» в маркетинговых сетях по сбыту тех или иных товаров. Дальнейшие примеры сетевых структур в биосистемах приведены в секции «Варианты сетевой организации».
Сетевые структуры в технических информационных системах. Примерами децентрализованных сетевых структур являются современные информационные сети, в особенности созданные на базе Интернета. Интернет, чьими предшественниками были Usenet, LISTSERV и особенно APRANET (в ведении Пентагона), с самого начала воплощал в себе сетевую архитектуру, которая не могла контролироваться из некоего центра и состояла из тысяч автономных компьютерных сетей, имевших многочисленные пути связи в обход электронных препятствий [5, 42].
В плане настоящей работы следует уделить внимание не только Интернету как физической сети из компьютеров, соединенных оптоволокном и другими связями, способными передавать данные [71], но и тем сетям пользователей, которые входят в состав системы World Wide Web. Система всемирной паутины была изобретена в 1980-е годы научными сотрудниками лаборатории физики высоких энергий CERN в Женеве как средство обмена информацией между ними и между их коллегами, однако вскоре стало ясно, что её потенциальные возможности существенно шире [72].
В эпоху Интернета сам термин «социальные сети» (social networks) приобретает узкое значение — это сообщества Интернет-пользователей, такие как глобальные сети Facebook, Bebo, Twitter, MySpace, LinkedIn, Nexopia (в первую очередь Канада), Tuenti (в основном Испания), «Одноклассники» (главным образом Россия и Украина), QQ (Китай) и др. Каждый пользователь со своим профилем представляет автономный узел такой сетевой структуры, способный избирательно устанавливать связи с другими пользователями («друзьями»). Во многих сетях чётко выражено и отмеченное выше объединительное начало — например, в специализированных сетях типа TheSocialGolfer или Great Cooks Community.
Тем не менее, несмотря на своё название, Интернет представляет собой смешанное полиструктурное явление. Сетевые структуры Интернета переходят — в силу внутренних и внешних для сети причин — в структуры иных организационных типов — иерархические (во многом в силу воздействия государственного аппарата, полицейских инстанций, образовательных учреждений) и рыночные (в результате коммерциализации Интернета).
Разумеется, всё сказанное об Интернете распространимо и на другие информационные сети, в частности, объединяющие пользователей мобильных телефонов (например, Facebook Mobile).
Нельзя не подчеркнуть, что «реальные» сетевые структуры из людей и их групп (организаций, фирм и др.) в социуме в возрастающей мере используют новые информационно-коммуникативные технологии, связанные с Интернетом, и приобретают тем самым всё более существенную виртуальную составляющую. Возможен — и достаточно важен в наши дни — обратный процесс: превращение первоначально виртуальных сетевых структур в реальные сетевые объединения людей, что часто имеет серьёзные социальные или политические последствия. Такая «девиртуализация» сетевых структур может происходить постепенно, но она может быть осуществлена и буквально в одночасье в результате акции флешмоба в виртуальной сети, как это имело место в ходе недавних революций в странах Северной Африки в2011 г. и на рубеже 2012-2013 гг. в Украине.
Плоские и объёмные сети. В структурном плане сеть отличается, как уже указывалось выше, отсутствием централизованной иерархии, так что отношения доминирования-подчинения между её элементами менее важны, чем горизонтальные кооперативные взаимодействия. Тем не менее, мы уже видели, что лидерство всё же возможно во многих сетевых структурах. В этой связи возникает основание для классификации сетевых структур.
Трансформация плоской сети в объёмную по мере роста сетевой структуры описана на примерах международных сетевых структур, борющихся за соблюдение прав человека, в частности, за права заключённых, особенно «узников совести» (сетевая структура Amnesty International). По мере увеличения масштаба сети, распределение связей между узлами (активистами, их группами) всё более соответствует принципу: в сети большинство узлов имеет малое число связей с другими узлами, и лишь меньшая часть узлов связано со многими другими узлами; этот принцип соответствует так называемым безмасштабным сетям, рассмотренным в ряде работ А.Л. Барабаши [30, 33]. Итак, немногие из её узлов концентрируют в себе социальную власть (social power, [60]) в рамках сетевой структуры. Это происходит вследствие того, что новые члены преимущественно присоединяются к узлам, уже связанным с другими узлами [60]. Аналогично, в социумах муравьёв коллективная активность, например, закладка муравейника, начинается сразу в нескольких точках несколькими «проектными командами». Впоследствии вновь прибывающие на стройплощадку рабочие муравьи присоединяются к тем «командам», где работа и так идёт наиболее успешно [7] – принцип стигмэргии, постулированный в классических работах П. Грассе.
Отметим, что в сетевом грядущем обществе, описанном в книге «Нетократия…» А. Барда и А. Зодерквиста [1], фактически предусмотрено формирование объёмных сетей, в которых выделяются достаточно властные лидеры («нетократы»). Правда, доступ к нетократам (кураторам сети) на вершине иерархизирующейся сетевой структуры затруднён системой секретных паролей, и эта особенность нетократии напоминает о масонских ложах Средневековья.
Объёмные сетевые структуры имеют аналоги в биосистемах, состоящих из одноклеточных организмов. Популяция клеток слизистого гриба (миксомицета) Dictyostelium discoideum на определенном этапе развития представляет собой компактную организованную клеточную массу — мигрирующий слизевик (псевдоплазмодий) [52, 70]. Это — типично сетевая структура с децентрализованной регуляцией. Под влиянием выделяемого многими клетками слизевика управляющего химического сигнала — циклического аденозиномонофосфата (цАМФ) – мигрирующий слизевик формирует плодовое тело в виде «грибка» с ножкой и шляпкой (где образуются споры). Неизменно плодовое тело формируется на верхней стороне мигрирующего слизевика, так как там концентрация цАМФ (и других химических факторов) максимальная. Клетки в этой зоне приобретают роль временных «проектных» (используя социальную терминологию) лидеров.
Несмотря на все возможные модификации, отсутствие единого фиксированного центра активности (начальника, босса, монопольного лидера) и в то же время кооперативные отношения между элементами (членами, участниками) отличают сетевые структуры, даже объёмные, от иерархических структур в биосистемах и человеческом социуме, на которых мы остановимся ниже.
ВАРИАНТЫ СЕТЕВОЙ ОРГАНИЗАЦИИ
Cетевые структуры в рассмотренном в настоящей работе значении децентрализованных кооперативных систем существенно различаются между собой по конкретным вариантам сетевой организации, которые могут реализоваться и в биосистемах, и в человеческом социуме, и в технических устройствах. Необходимо особо оговорить, что, если большинство живых существ просто живёт в сетевых структурах различных подтипов, то человек обладает способностью изменять, творить эти структуры, сообразуя со своими интересами, целями, потребностями. В отличие от структур, формируемых другими живыми существами, в структурах человеческого социума и в созданных человеческим разумом технических системах могут стихийно или сознательно комбинироваться элементы разных «биологических» вариантов; они могут дополняться уникальными «человеческими» организационными сценариями.
Какие варианты сетевой организации могут быть плодотворно экстраполированы от биологических систем на человеческий социум и технические информационные устройства? В недавних публикациях [19-21] мы рассмотрели шесть вариантов: клеточные, эквипотенциальные, модульные, эусоциальные, нейронные и эгалитарные сети.
Нейронные сети. Данный вариант сетевой организаии, как показывает само название, присущ системам, содержащим нейроны (нервные клетки), то есть структурам нервной системы, в особенности мозгу. В технических устройствах эффективно работают их аналоги (искусственные нейронные сети), такие, как перцептрон, сеть Хопфилда, слой Кохонена и др. Все эти сети являются объемными – сочетают в себе сетевой и иерархический принцип – и обладают рядом общих свойств, в том числе:
Многие социально-технологические проекты, имеющие в виду создание сетевых структур, фактически руководствуются принципами нейронного организационного варианта. Уроки в школе/вузе могут ориентироваться на нейронные сети как полезную организационную модель. Это способствует параллельной обработке информации творческими подгруппами в рамках сети, составленной из учеников класса или студентов группы. Сеть в целом в этом случае ассоциативно строит образ целого решения поставленной задачи на базе фрагментов, предложенных каждой из творческих подгрупп («слоёв» коллективной «нейронной сети»). Доминантой в ходе самостоятельных работ школьников/студентов по «нейронному» сетевому сценарию должно быть самообучение как каждого индивида, так и на групповом уровне – уровне сети как коллективного «мозга». Рис. 3. Сетевая структура у микроорганизмов. Сканирующая электронная микрофотография клеток кишечной палочки Escherichia coli M-17 (слева) и дизентерийной палочки Shigella flexneri (справа), видны контакты между клетками, формирующими кластеры (сеть с высоким уровнем кластеризации, см текст ниже). Дар О. Рыбальченко.
Рис. 3. Сетевая структура у микроорганизмов. Сканирующая электронная микрофотография клеток кишечной палочки Escherichia coli M-17 (слева) и дизентерийной палочки Shigella flexneri (справа), видны контакты между клетками, формирующими кластеры (сеть с высоким уровнем кластеризации, см текст ниже). Дар О. Рыбальченко.
Их примером является «чувство кворума» (quorum sensing) [17, 55, 89] — «демократический» механизм, обусловливающий координированные действия элементов микробной сетевой структуры. Согласованное групповое поведение при этом основано на своеобразном химическом «голосовании». Бактерии в популяции выделяют сигнальное вещество, и когда концентрация этого вещества в окружающей среде достигает порогового значения, все клетки меняют свое поведение (например, начинают активно агрегировать с формированием микроколоний). На молекулярном уровне изменение поведения микробов обеспечивается изменением уровня активности определенных генов в ответ на связывания феромонов с рецепторами.
Не только нейронные сети состоят из клеток (или их технических или социальных аналогов). Сетевые структуры формируются и многими другими типами клеток – как свободноживущими (микроорганизмами), так и находяцимися в составе тканей многоклеточных организмов. Некоторые из таких клеточных сетей не имеют даже частичных лидеров, т.е. в нашей классификации представляют плоские, а не объёмные сетевые структуры. Помимо этого, в отсутствие лидеров координация поведения отдельных клеток в интересах работы всей сети во многом обеспечивается контактами между клетками (рис. 3), а также дистантными (охватывающими всю структуру в целом) химическими коммуникационными сигналами.
Модульные сети присущи биосистемам, построенным из повторяющихся структурных единиц (модулей); они также отличаются преобладанием плоской (безлидерной) сетевой организации. Пример представляют колониальные кишечнополостные, чьё тело состоит из связанных между собой единым стеблем (ценосарком) полипов или медуз. Такая колония представляет собой децентрализованную структуру. Каждый полип, совершая те или иные действия (например, сжимаясь и создавая ток жидкости) слабо воздействует на целую систему, но его эффект усиливается, если поведение этого полипа соответствует поведению большинства остальных полипов в системе. «Если в момент сжатия пульсара, (т.е. совершающего периодические сжатия и растяжения полипа – примечание О.А.) в него поступает мощное течение… со стороны других пульсаров, также испытывающих сокращение в это же время, то побеждает группа консолидированных пульсаров, а у остальных происходит задержка собственного сжатия, после которой происходит сдвиг фазы пульсаций» [12].
Аналогичный принцип в сетевых структурах в человеческом социуме именуется «сетевой рычаг» — «такая организация взаимодействия между членами сети, при которой каждый из них, предоставляя для формирования корпоративного ресурса часть своего личного ресурса…, в случае необходимости получает в собственное распоряжение на порядок больший объём консолидированного корпоративного ресурса» [28]. Сетевой рычаг может быть статусным, информационным, комунникационным, ресурсным, в зависимости от того, что именно объединяется и совместно используется звеньями сетевой структуры.
Эквипотенциальные сети отличаются совершенно плоским характером [22, 24]. Примером служат стаи многих рыб или морских беспозвоночных. В отсутствие лидера первой в движущейся стае плывёт случайная особь, вскоре сменяемая другой особью. У молоди сайды одна и та же рыба «лидерствует» «от долей секунды (0.25—0.5) до нескольких секунд…, после чего скатывается в середину или даже в самый арьергард стаи» [24]. Функционирование такой сети во многом определяется взаимодействием между соседями, при которым информация в виде зрительных или механических (течение воды, воспринимаемое боковой линией рыб) передается по эстафете. Эстафетная передача сигналов позволяет стае совершать сложные координированные маневры, например, рассыпаться при приближении хищника или формировать серповидную структуру, зажимая стаю более мелких рыб-жертв между рогами этого «полумесяца». С точки зрения моделирования принципов функционирования эквипотенциальных сетей в технических устройствах представляют интерес работы по алгоритмизации поведения их элементов.
Предложена модель, где поведение стаи рыб алгоритмизировано на основе следующих норм поведения особей: 1) уходить от слишком близко располагающихся соседей; 2) двигаться в том же направлении, что и соседи; 3) не допускать слишком большого расстояния между данным индивидом и соседями. Получатся три окружающих индивида зоны — 1 – отталкивания, 2 – следования за соседями и 3 – притяжения к ним. Сохраняя постоянными зоны отталкивания и притяжения и варьируя размер зоны следования 2, можно получить различные геометрические образы на уровне целой стаи — разреженную, стоящую на месте стаю; тор («бублик»), где особи двигаются вокруг центра масс; ходовую стаю с параллельным движением особей [44].
 Рис. 4. Формирование «рабочей команды» в колонии муравьёв Camponotus socius. Ситуационный лидер (крупный муравей слева) рекрутирует членов команды, используя химические коммуникационные сигналы. Схема основана на рисунке из книги [58] (где схема обозначена как Fig. 6-24).
Рис. 4. Формирование «рабочей команды» в колонии муравьёв Camponotus socius. Ситуационный лидер (крупный муравей слева) рекрутирует членов команды, используя химические коммуникационные сигналы. Схема основана на рисунке из книги [58] (где схема обозначена как Fig. 6-24).
Аналогами эквипотенциальных сетей в человеческом обществе являются безлидерные структуры, спонтанно формирующиеся в экстремальных ситуациях (например, в обстановке мятежа типа «майдана»). Индивиды при угрожающих жизни ситуациях, могут следовать достаточно примитивным правилам поведения, напоминающим алгоритмы поведения рыб в стае, например: 1) поиск кратчайшего пути к безопасному месту; 2) избегание столкновений с соседями; 3) движение в том же направлении, что и другие [56]. Получается достаточно сложное результирующее поведение толпы. Причём, к сожалению, указанное поведение индивидов способствует блокировке движения всей толпы и снижает вероятность спасения людей в ней [84].
Эусоциальные сети являются объёмными, т.е. включают в себя частичных лидеров. Так, в эусоциальных структурах общественных насекомых, в частности, муравьёв, есть «рабочие команды» (кланы), возглавляемые ситуационными лидерами и выполняющие задачи типа рытья почвы или сбора выделений тлей [3, 4, 25, 58] (рис. 4). Но лидеры кланов выступают как частичные лидеры в рамках децентрализованной структуры более высокого порядка (колонны, плеяды)2. Обращает на себя внимание также наличие в составе социума насекомых, в частности, муравьёв, неспециализированных, не задействованных рабочими командами особей. Такой пул неспециализированных и часто малоактивных особей [79] представляет резерв колонии и может быть мобилизован для решения особо важных задач.
В человеческом социуме аналогичным образом специализированный частичный лидер и помощники-эксперты взаимодействуют с неспециализированными членами сетевой структуры хирамы, которые во многих хирамах численно преобладают; сходный принцип специализированного меньшинства и поддерживающего неспециализированного большинства характерен для многих клубов по интересам и в то же время для ряда «тёмных» — криминальных и террористических сетей. Особенности сетевой организации общественных насекомых используются при создании алгоритмов искусственных систем «роевого интеллекта» (Swarm Intelligence) с распределённым контролем и оптимизацией, причем построенные по образцу насекомых технические системы отличаются значительной гибкостью и стабильностью функционирования в динамической среде.
Эгалитарные сети также являются объёмными. Они характерны для некоторых приматов и отличаются следующими принципами:
Отметим, что по сходным принципам были организованны не только многие первобытные общности охотников-собирателей, но и современные творческие лаборатории, особенно включающие в себя выдающихся учёных, где каждый волен заниматься своей индивидуальной тематикой и разрабатывать собственные концепции; эту свободу сетевая структура ограничивает лишь временными, частными обязательствами, обусловленными совместными проектами, публикациями, конференциями.
КОЛИЧЕСТВЕННЫЕ ХАРАКТЕРИСТИКИ СЕТЕВЫХ СТРУКТУР: КРИТЕРИИ ЦЕНТРАЛИЗАЦИИ (ИЕРАРХИЗАЦИИ)
В последние десятилетия в литературе появилось значительное количество исследований, посвященных количественным параметрам, позволяющим оценивать свойства сетевых структур [30, 33? 39, 54, 71-73, 80]. Некоторые из количественных характеристик сетевых структур фактически позволяют нам различать сети в узком смысле, иерархии и (квази)рыночные структуры. Под этим углом зрения рассмотрим в краткой форме ряд важных сетевых свойств, вычлененных представителями современной «network science».
Например, понятие геодезической линии (geodesic line), или длины пути (path length), соответствует минимальному количеству узлов, которые надо пересечь, чтобы пройти расстояние между заданной парой узлов в сети [72].
Особый интерес представляют критерии, которые позволяют нам выяснить, в какой мере данный узел (или группа узлов) занимает центральное положение по отношению к другим узлам в сетевой структуре. Речь идёт о нескольких параметрах (степень узла, его близость, промежуточность, величина собственного вектора3), которые могут быть применены как для описания степени центральности отдельного узла, так и для характеристики всей сети или её частей (подсетей, кластеров), превращаясь в этом случае в критерии их централизации (иерархизации).
Степень узла (node degree). Степень узла в сети может быть обозначена буквой К; она соответсвует количеству непосредственных соседей данного узла – числу других узлов, с которым данный узел соединён прямыми связями. Например, узел I на рис. 5 имеет К = 5. Синоним термина «степень узла» – термин степенная центральность (degree centrality). Во многих публикациях величину К нормируют для каждого узла в сети, деля её на суммарное число связей в данной сети; полученная нормированная степень узла варьирует от 0 до 1 [53]. Примером работ, где использовалась степень узла К, являются статьи, в которых выясняли относительную важность каждого из членов сети, совершившей террористические акты 11 сентября2001 г. в США [62, 63].
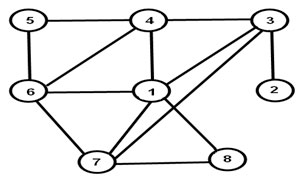
Рис. 5. Простая сеть, в которой узлы различаются по числу непосредственных соседей – по «степенной центральности», или просто «степени узла».
С этих позиций может быть предложена мера централизации (иерархизации) всей сети: она определяется как различие в параметре К у разных узлов одной и той же сети. В сети на рис. 5 узел 1 имеет максимальное количество (Кmax) связей с другими узлами (5), а узел 2 — минимальное количество (Кmin) связей (1).
В централизованной сети (т. е. иерархии) большая часть связей сконцентрирована вокруг одного или немногих узлов, а в децентрализованной сети (сети в узком смысле, который мы используем в дальнейшем тексте работы) разница между числом связей у разных элементов сети мала. В строго количественном смысле централизацию всей сети (<K>) с точки зрения разницы степеней её узлов оценивают по формуле: <K> = Sni=1 (Kmax– Ki)/<Kmax>, где Ki– степень каждого из узлов (i) в данной сети, а <Kmax> – максимально возможное значение централизации при заданном числе узлов сети. «Сетевая наука» утверждает, что макисмальная централизация достигается, если узлы в сети соединены по сценарию «звезды» («звёздный граф», рисунок); <Kmax> подсчитывается по формуле <Kmax> = n2 - 3n + 2, где n есть число узлов в данной сети [53]. Централизация (иерархизация) сети в виде «звезды» равна единице.
Степень посредничества (betweenness degree). Этот критерий центральности узла в сети основан на числе геодезических линий («кратчайших путей»), которые проходят через данный узел и связывают пары узлов в той же сети [53, 72]. В сети на рис. 5 через узел 1 проходит максимальное количество (Вmax) путей, связывающих другие узлы (10 путей — это пути 318, 317, 316, 314, 418, 417, 416, 617, 618, 7184), а через узел 2 — минимальное количество (Вmin) путей (05). Примечательно, что некоторые узлы сети с низкой степенной центральностью тем не менее могут характеризоваться высоком уровнем степени посредничества, также обозначаемой как посредническая центральность узла (betweenness centrality). Существуют сети, состоящие из подсетей, связанных между собой лишь одним узлом (cut point node), через который проходит много связывающих другие узлы путей; такой узел отличается высокой посреднической центральностью. Таковы террористические трансграничные сети, например, Аль-Каида, имеющая локальные подсети [41, 62, 63]. Бактериальные биоплёнки, в том числе формируемые нитчатыми цианобактериями, нередко имеют сегментированный характер: центральная колония образует периферические колонии («отстрелы», [23, 26]), связанные с центральной колонией лишь узкими перешейками, которые приобретают особое значение для сохранения целостности структуры. Высокий уровень посреднической центральности был характерен для малой группы дельфинов, находившихся на границе между двумя плотными популяциями дельфинов в бухте Даутфул Саунд (Новая Зеландия) [65]. Сегментированный (фрагментированный) характер имеют и большие стаи рыб [45-47, 91].
Централизация (иерархизация) всей сети в рамках критерия «посредническая центральность узлов» оценивается, исходя из разности между максимальным и минимальным значениями степени посредничества у узлов одной сети, а узел с максимальной степенью посредничества оказывается лидером (центральным звеном). В децентрализованных сетевых структурах указанная разность близка к нулю. Никто из членов сети не может похвастаться наибольшим числом связей с товарищами по сети.
Количественно, аналогично степенной централизации, посредническая централизация (иерархизация) сетевой структуры <B> подсчитывается как сумма разностей между Bmax , максимальным значением посреднической центральности, и Bi, посредническими центральностями всех узлов сети. Полученная сумма нормируется делением на максимально возможную сумму разностей <Bmax>, которая также наблюдается в случае «звезды» и подсчитывается по формуле:
<B> = Sni=1 (Bmax – Bi)/ n3 - 4n2 + 5n - 2.
Степень близости (closeness centrality). Данный параметр (Б) возрастает по мере сокращения средней длины пути от данного узла до других узлов. Один из способов подсчёта: находится обратная величина усредненной длины кратчайших путей между данным узлом и другими узлами той же сети: Б = 1/(Sni=1 li/n-1), где l – длина кратчайшего пути между данным узлом и каким-либо другим узлом i той же сети; n – число узлов в сети. В централизованной сети (иерархии) центральное положение одного или немногих узлов определяет сравнительно короткие пути между ними и прочими элементами, т. е. сравнительно высокое значение Б, а в децентрализованной сети разница между длиной путей, исходящих от разных элементов сети, мала, и степень централизации, рассчитанная по разности в степени близости (closeness centrality), близка к нулю.
Централизация (иерархизация) сети с точки зрения «близостной центральности» – величину <Б> – определяют аналогично степенной и посреднической централизации как сумму разностей между максимальной величиной близости узла в этой сети и величинами близости, присущими другим узлам той же сетевой структуры; полученную степень иерархизации сети нормируют, деля её на (n2+ 3n -2)/(2n-3) [53].
Степень по собственному вектору (eigenvector centrality). Дополнительный критерий центральности узла в сети – и, при распространение на всю сетевую структуру – её централизации (иерархизации) определяется, исходя из числа влиятельных (т.е. связанных со многими узлами) соседей данного узла. Иначе, говоря, в отличие от простой степени узла, не просто подсчитывается количество непосредственных соседей, но учитывается их «вес» в сетевой структуре. Данный параметр может быть большим у данного узла или потому, что узел имеет много связей, или потому, что он связан, пусть с немногими, но «весомыми» соседями по сети. Индивид в социальной сети может иметь высокую степень по собственному вектору или потому, что он знаком со многими людьми (которые сами по себе не столь важны), или потому, что знает немного людей, но они занимают важные позиции в обществе [72]. Подсчитывая сумму разностей между максимальной величиной степени по собственному вектору и таковыми других узлов той же сети, мы можем определить степень централизации всей сетевой структуры по собственным векторам её узлов.
ПРИМЕНЕНИЕ КРИТЕРИЕВ ЦЕНТРАЛИЗАЦИИ ДЛЯ РАЗГРАНИЧЕНИЯ СЕТЕЙ (В УЗКОМ СМЫСЛЕ) И ИЕРАРХИЙ
Каждый из четырёх предложенных выше критериев даёт свою оценку степени централизации (иерархизации) структуры. В цитированной классической работе Фриман [53] допускал, что разные критерии централизации соответствуют разным иерархиям, формирующимся в той же сети. Так, узел с максимальным значением посреднической центральности (скажем, связной между двумя локальными кластерами сети Аль-Каеды) может иметь мало непосредственных контактов (узлов-соседей) и потому сравнительно низкую величину степенной центральности. Несмотря на это, есть и замечательные корреляции между разными оценками степени иерархизации структуры.
Они в наибольшей мере характерны для двух крайних случаев [53] (рис. 6): 1) максимально централизованные структуры, примером которых служит граф в форме звезды, для которого характерны высокие значения централизации (иерархизации) по всем рассмотренным выше критериям (рис. 6, слева); в «звезде» один и тот же центральный узел характеризуется максимальной степенной, близостной и посреднической центральностью, а также центральностью по критерию собственного вектора; 2) полностью однородные сетевые структуры, в которых присутствуют все логически возможные связи между узлами (это так называемые полные графы), а также геометрически правильные решётки.
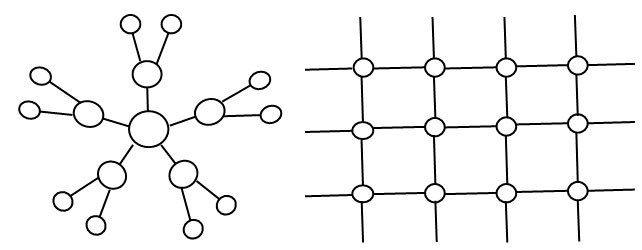
Рис. 6. Иерархическая структура («звезда»), в которой центральный узел имеет максимальные значения центральности по всем 4 критериям, рассматриваемым в данной статье (слева); полностью плоская сеть (решётка), где все узлы имеют одинаковые значения центральности по всем критериям (справа).
Все их узлы имеют одну и ту же степень центральности по четырём указанным выше критериям. Поэтому вся сеть имеет нулевую централизацию по всем данным критериям (рис. 6, справа). Эти критерии центральности узлов и иерархизации всей структуры дают существенно расходящиеся результаты в структурах с промежуточными характеристиками.
Именно такие характеристики наблюдаются у объёмных сетей, которые широко распространены в мире биологических, социальных и технических информационных систем. Их децентрализованная, распределённая организация с наличием множественных частичных лидеров соответствует тому факту, что разные узлы оказываются максимально центральными по разным критериям. Мы не случайно кратко рассказали о сетевой структуре типа хирамы выше. Очевидно, что в хираме разные типы частичных лидеров могут иметь высокий ранг по разным критериям центральности. Внешний лидер, который должен выполнять функции связного и брокера, будет скорее всего иметь высокий уровень посреднической центральности. Психологический лидер должен отличаться высоким уровнем степенной центральности, ибо в его функции входит непосредственное общение с большим числом членов сети. Организационный лидер должен без труда передавать сообщения другим «хирамщикам», которых он и должен организовать, т.е. следует ожидать, что он имеет высокий уровень близостной центральности.
В противоположность такой распределенной сети, жёсткая иерархия стремится к близким значениям центральности узлов и иерархизации всей структуры – по всем четырём критериям. Один и тот же узел или малая подгруппа узлов оказывается наиболее центральной и по степени узлов, и по их близости, и по их посреднической ценральности, и по собственному вектору. В то же время, большинство прочих узлов той же структуры, находящихся в подчинённом положении, отличаются существенно более низкими значениями центральности по всем критериям. Да будет мне позволено привести политический пример. И. В. Сталин как политический лидер контролировал потоки информации в пределах всего аппарата управления (отличался максимальной посреднической центральностью), лично общался с колоссальным числом подчинённых (т.е. имел наивысшую степенную центральность) и, как правило, без всякого труда доводил свои распоряжения даже до тех граждан, с кем не имел прямого контакта, используя все имевшиеся в ту эпоху средства массовой информации (поэтому его близостная центральность также, очевидно, была максимальной во всей структуре).
Можно также привести пример из жизни Аль-Каеды. В ряде работ проанализирована структура групп террористов-смертников, разрушивших башни ВТО и Пентагон 11 сентября2001 г. Из полученных данных следует однозначный вывод, что, несмотря на децентрализованный во многом характер всей международной сети Аль-Каеды, эти группы носили иерархический характер. Их главарь Мохаммед Атта, лично участвовавший в угоне самолёта, врезавшегося в северную башню ВТО, имел, из всех членов террористической группы, максимальный уровень степенной и близостной центральности; он был вторым во всей группе по уровню посреднической центральности [62, 63].
Итак, тенденция к совпадению вершин иерархий, построенных по разным критериям центральности, говорит о том, что вся структура есть не сеть (в узком смысле), а централизованная иерархия. Наличие этой тенденции позволяет нам отличить истинную централизованную иерархию от распределённой сети, пусть даже и объёмной, которая имеет частичных ситуационных лидеров.
Отметим, что рассмотренные нами критерии центральности узлов сети позволяют также дифференцировать объёмные и плоские сетевые структуры. Рассмотрим это на примере степени узла (степенной центральности) К.
Количественной характеристикой целой сети является величина p, вероятность того, что произвольный узел сети имеет заданную степень k; альтернативная характеристика — интегрированная величина P, т.е., вероятность того, что степень узла превышает заданное значение.
Плоские сети (лишённые даже частичных лидеров) типа упомянутых стай рыб могут быть представлены как хаотические рэндом-сети создаваются по алгоритму «берем п узлов и с вероятностью р проводим соединительную прямую (ребро) между ними» [30]. Для таких сетей, впервые проанализированных Эрдёс и Реньи и названных по первым буквам фамилий ER-сети, характерно нормальное (пуассоновское) распределение вероятностей для значений k. Иначе говоря, ER-сети включают так называемые типовые узлы, чьи степени имеют максимальную вероятность (вершину гауссовой кривой).
Однако многие реальные структуры ведут себя в соответствии с гиперболической зависимостью (power law) p от k: p(k) = a × kg, где a — константа, а g < 0. Это безмасштабные сети (scale-free networks), исследованные в недавних работах Барабаши и Альберта [30]. Безмасштабная сеть строится так: «… сети растут путем добавления новых узлов, соединяемых ребрами с уже существующими узлами… Новый узел с бóльшей вероятностью присоединяется к узлу с бóльшим количеством связей» [30]. Примеры таких сетей можно найти в Интернете, где новые пользователи предпочтительно вступают в связь с узлами, и так имеющими большее количество связей. Безмасштабные сети содержат малое число хабов (hubs) – частичных лидеров с многочисленными связями; численно преобладают узлы с небольшим числом связей.
Итак, гиперболический (или близкий к нему) тип зависимости вероятность того, что произвольный узел сети имеет заданную степень К, от значения K позволяет нам дифференцировать объёмные сети с хабами (частичными лидерами) от плоских сетей без частичных лидеров, которым присуще пуассоновское распределение p(K) ~ K.
Следует подчеркнуть, что рассмотренный критерий распределения p(K) не позволяет различить объёмные сети и централизованные иерархии когда среди хабов выделен один центральный узел (пейсмейкер, босс, лидер, доминант), подчиняющий себе остальные узлы, располагающиеся на разных ступенях иерархической статусной лестницы.
РАЗГРАНИЧЕНИЕ СЕТЕЙ И (КВАЗИ)РЫНОЧНЫХ СТРУКТУР
Следует ещё раз подчеркнуть, что (квази)рыночные структуры – форма организации систем в человеческом обществе, в основе которой автономия элементов, обмен на эквивалентной основе и конкурентные отношения. Для некоторых биосистем также характерны в основном конкурентные (а не кооперативные) взаимоотношений между автономными единицами, что заставляет вспомнить о рынке в человеческом обществе. Как разграничить сетевые (в строгом смысле) и (квази)рыночные структуры?
В этом случае, как представляется, можно использовать не столько математический аппарат (см., однако, конец этого подраздела о кластеризации), сколько экологические критерии.
В экологических терминах отношения между узлами в сети (скажем, особями в группе животных) могут быть позитивными (полезными) или негативными (вредными). Позитивные, или плюс-отношения между особями повышают их шансы на выживание и эффективность размножения, усиливают приспособляемость (fitness). Такие отношения обозначаются в экологии как кооперативные (мутуалистические), и системы, где они преобладают, обозначаются как сетевые структуры в строгом смысле, при дополнительном условии отсутствия централизованной иерархии в них. Хорошо известна симбиотическая структура, образуемая грибом и водорослью – лишайник.
Негативные, или минус-отношения снижают выживаемость, эффективность размножения, адаптивность. Если они преобладают в системе, перед нами в целом конкурентная, квазирыночная, структура. Не случайно, что с экологической точки зрения слово «конкуренция» во многом синонимично слову «антагонизм», хотя они различаются нюансами значений.
Аналогично рыночным структурам человеческого общества, многие животные, в том числе насекомые, ракообразные и др., формируют скопления, где они продолжают в большой мере вести себя независимо и скорее конкурируют, чем кооперируют друг с другом. Так, насекомые типа луговых кобылок формируют огромные анонимные стаи, известные как саранча. Если некоторые индивиды в этой квазирыночной структуре побеждают в конкурентной борьбе остальных, то проигравшие оказываются на положении подчиненных особей, а вся структура из квазирыночной превращается в иерархическую. Альтернативный исход: полное уничтожение проигравших в конкурентной борьбе (банкротство в коммерческой сфере; гибель бактериальных клеток под воздействием антибиотиков, выделенных бактериями-конкурентами и другие примеры).
Данный критерий разграничения сетей и (квази)рынков связан с концепцией так называемых сетей со знаками (или графов со знаками), которые могут иметь положительные или отрицательные рёбра/связи между узлами. За пределами экологии сети со знаками находят применение при исследовании социальных структур. «Например, в сети знакомств мы можем определить дружбу как положительную связь и вражду как отрицательную связь» [72].
Следует отметить, что антагонистические или конкурентные отношения могут оказаться полезными для взаимодействующих элементов структуры, хотя и представляют собой минус-отношения по непосредственному эффекту (так, конкуренция за кислород между двумя плодами-близнецами в одной матке ограничивает его потребление каждым плодом; конкуренция между двумя фирмами за потенциальных потребителей снижает шансы успешного маркетинга для каждой из конкурирующих фирм). Тем не менее, хорошо известно, что положительные эффекты конкуренции связаны с компенсаторными усилиями по преодолению конкурентов. Всё это в конечном счёте связано с тем, что Адам Смит обозначал как «невидимая рука» рынка (которая имеет аналоги и в биосистемах). Известный пример – стимулирование иммунной системы организма потециально вредоносными бактериями, которые образуют недостаточные для серьёзного ущерба дозы токсичных веществ; они мобилизуют иммунную систему, заставляя её «держать порох сухим».
Сетевые структуры, где кооперация элементов доминирует над конкуренцией между ними, часто явно отличаются от (квази)рыночных структур по такому параметру как коэффициент кластеризации (С) узла в сети. Коэффициент С соответствует отношению реального числа ребер между его соседями и их потенциально возможного числа. Для узла i Ci= Ei/[ki(ki-1)/2], где Eiреальное число ребер, ki – степень узла, а в знаменателе записано суммарное число потенциально возможных ребер между непосредственными соседями узла i (при котором сеть или её часть превращается в полный граф). Коэффициент кластеризации может быть усреднен для любой части сети или для сети в целом, становясь ее интегральной характеристикой: C = 1/n ∑Ci. Если сеть включает в себя части с тесными связями между узлами, т.е. с высоким значением C, которые соединены лишь редкими связями между собой, то такая сеть отличается наличием клик, или «коммунальной структурой» (community structure, cliquishness). Исходно социальный термин «клика» переопределяется в теории сетей как «набор узлов, где каждый узел прямо связан со всеми остальными» [91].
Сетевая структура, построенная в основном на кооперации её узлов, часто состоит из большого числа плотных подсетей (кластеров), разделённых зонами с редкими связями (примером могут служить именно так устроенные стаи многих видов рыб). Такая кластеризация в существенно меньшей мере присуща традиционным рынкам в человеческом обществе и аналогичным конкурентным системам в биологическом мире. Соответственно, кооперативные сетевые структуры оказываются гетерогенными в плане коэффициента кластеризации. Коэффициент С достигает высокого уровня у тех узлов, которые включены в состак кластеров (клик, подсетей); этот коэффициент низок в зонах сети, расположенных между её кластерами. Рынки и их аналоги характеризуются более равномерно распределёнными по всей рыночной структуре значениями С , что связано с автономных характером каждого из узлов – каждого из агентов, вступающих в (квази)рыночные отношения (контракты; метаболические отношения в биосистемах).
Биологическая, социальная или техническая система могут эволюционировать во времени; они могут превращаться из горизонтальных структур в иерархические или из кооперативных в конкурентные (или наоборот). Однако, тот факт, что структурные взаимопревращения возможны, не означает, что границы между сетями, иерархиями и (квази)рынками вообще не существуют. Переходы между разными структурами связаны с существенными организационными изменениями; они могут намеренно реализоваться в человеческом обществе с целью создания новых возможностей для приспособления к изменяющимся внешним условиям. То, что некоторые виды обезьян могут формировать иерархии при коллективной охоте и превращать их в более эгалитарные структуры сетевого типа на отдыхе, не значит, что не наблюдается чёткого различия между данными структурами.
Таким образом, узкое понимание термина «сети», противопоставляющее их иерархическим и (квази)рыночным структурам, оказывается распространимым на системы, не относящиеся к человеческому социуму, в частности, на разнообразные биосистемы. Мы предлагаем математические и экологические критерии для разграничения по линиям сеть-иерархия и сеть-(квази)рыночная структура.
Не приравнивая человеческие социальные системы к биосоциальным системам других форм живого, мы можем констатировать, что сравнительный анализ организационных вариантов децентрализованных распределённых кооперативных сетевых структуры в биосистемам, человеческом социуме и технических информационных системах дает «пищу для ума» исследователям и практическим разработчикам социальных структур с научно-исследовательскими, педагогическими, политическими, военными (в том числе антитеррористическими) целями.
А.В. Олескин, кафедра общей экологии, биологический факультет МГУ;
директор Клуба «Биополитика» при МОИП